;Resize,width=767;)

L'apoptosi è un meccanismo di morte cellulare programmata con cui il nostro organismo elimina cellule infette, invecchiate, difettose o semplicemente non più necessarie. È un processo pulito, ordinato ed è fondamentale per garantire l’omeostasi e l’equilibrio dell’organismo. In effetti, quando Seneca nella Lettera 24 a Lucilio scriveva “Ogni giorno moriamo” (Cotidie morimur) non aveva tutti i torti. Una review del 2001 di Andrew G Renehan, Catherine Booth e Christopher S Potten, stima che nel nostro corpo, ogni giorno 10 miliardi di cellule decidono di morire (o ricevono un ordine di autodistruggersi). L’ordine di autodistruzione può arrivare sia dall’interno stesso della cellula che dall’esterno. A prescindere da chi dia l’ordine il risultato è lo stesso: un ordinato spacchettamento delle strutture cellulari che vengono poi “mangiate” dagli spazzini del sistema immunitario senza lasciare traccia.
Cos'è l'apoptosi e quali meccanismi cellulari coinvolge
L'apoptosi, termine che fu coniato nel 1972 da John Kerr, Andrew Wyllie e Alastair Currie, dal greco "cadere via", è un meccanismo di morte cellulare controllata, con cui l'organismo elimina cellule infette, invecchiate o difettose, come quelle tumorali. È molto diverso dal caos della necrosi causata da danni acuti alla cellula, come la rottura della membrana: l’apoptosi, infatti, è un processo ordinato, pulito e che richiede energia.
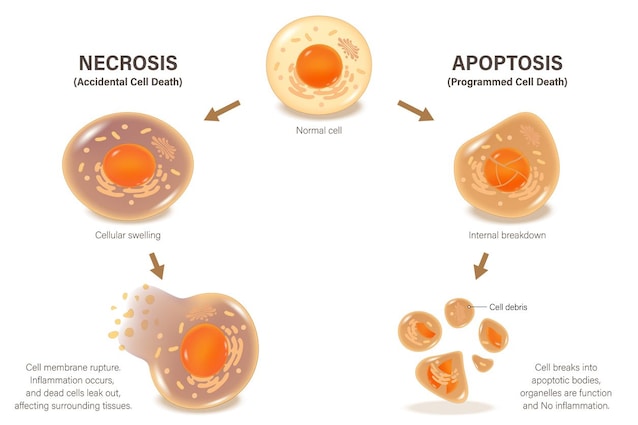
Questa degradazione può però partire da due punti di origine molto diversi: dall'esterno della cellula o dal suo interno. Una revisione pubblicata sul Biochemical Journal descrive con chiarezza questa distinzione: l'apoptosi viene attivata quando specifici recettori presenti sulla cellula vengono attivati (via estrinseca), oppure quando viene “aperta” la membrana esterna dei mitocondri presenti nella cellula (via intrinseca). Il punto d’arrivo è uguale per entrambe e coinvolge una famiglia di enzimi chiamati caspasi, che tagliano molecole e strutture cellulari come vere e proprie forbici molecolari.
La via estrinseca
Questa via si attiva quando segnali provenienti dall'esterno (ligandi) della cellula "ordinano" alla cellula bersaglio di autodistruggersi. I protagonisti sono i recettori di morte, una famiglia di proteine transmembrana e le coppie recettore di morte-ligando più studiate sono: TNF con il suo recettore TNFR1, Fas-L con il recettore Fas, e i recettori DR4 e DR5, con il ligando TRAIL. Questi recettori hanno una zona esterna alla cellula (dominio extracellulare) che riconosce il ligando e un dominio intracellulare chiamato "dominio di morte". Una volta attivato, il dominio di morte chiama a raccolta una serie di proteine formando un complesso multiproteico chiamato DISC (Death-Inducing Signaling Complex), che a sua volta recluta la caspasi 8.
A questo punto, o la caspasi 8 attiva un’altra caspasi (la 3) dando il via all’autodistruzione; oppure, se è insufficiente, si fa aiutare, attivando un passaggio della via intrinseca per amplificare l’ordine di esecuzione, come ben documentato da numerosi studi.
È usata non solo per combattere le infezioni, ma principalmente per mantenere in equilibrio il sistema immunitario, per esempio quando c’è bisogno di eliminare linfociti T non più necessari dopo un’infezione, o “impazziti” che attaccano cellule sane. O anche per mantenere il privilegio immunitario di organi come testicoli, occhi, follicoli piliferi. Queste aree sono una sorta di paradiso fiscale dove il sistema immunitario non può entrare: quando un linfocita cerca di imbucarsi, viene prontamente indotto all'autodistruzione tramite la via estrinseca.
La via intrinseca
In questo caso, il segnale di morte origina dall'interno della cellula, in risposta a danno al DNA, mancanza di fattori di crescita, carenza di ossigeno o accumulo di radicali liberi. Il fulcro molecolare di questa via sono i mitocondri e la famiglia di proteine BCL-2, descritte nel dettaglio dalla revisione del 2009 di Brunelle e Letai. In questa famiglia esistono due fazioni: una che vuole distruggere la cellula (proteine pro-apoptotiche) e l’altra che la vuole salvare (anti-apoptotiche). Conosciamole meglio:
- Bax e Bak, “i sicari”. Fanno partire di fatto l’autodistruzione;
- BCL-2 e BCL-xL “i poliziotti”. Tengono sotto controllo Bax e Bak, inattivandoli;
- Proteine BH3-only, i “sensori”, i cui nomi sembrano usciti da un programma per ragazzi di qualche (ahimè) decennio fa: Bid, Bim, Puma, Noxa, Bad. Percepiscono segnali di stress della cellula e corrono a far evadere Bax e Bak inibendo i poliziotti.
Quando Bax e Bak sono liberi, inducono la permeabilizzazione della membrana mitocondriale esterna (MOMP), creando un poro nel mitocondrio da cui esce il citocromo c che a sua volta si lega a una proteina, Apaf-1. Insieme formano un complesso degno della fusione tra Goku e Vegeta: l’apoptosoma, un “Gogeta” molecolare che attiva la caspasi 9.
Per evitare che altri protettori della cellula, come le proteine IAP (Inhibitors of Apoptosis Protein), si intromettano, dal mitocondrio esce anche la proteina SMAC/DIABLO che blocca le IAP facendo proseguire l’autodistruzione.
Bonus: il meccanismo perforina/granzima
Oltre alle due vie classiche, il meccanismo perforina/granzima, altrettanto importante, è utilizzato dai linfociti T citotossici (CD8+) e dalle cellule Natural Killer (NK) per eliminare cellule infette da virus o cellule tumorali, come riporta una revisione del 2007 pubblicata su Toxicologic Pathology. Quando incontrano e riconoscono una cellula bersaglio, rilasciano nello spazio intercellulare perforina (che forma pori nella membrana bersaglio) e granzima A e B. La granzima B taglia Bid e attiva le caspasi, mentre la granzima A agisce con un meccanismo caspasi-indipendente che porta comunque a danno del DNA. È un'arma rapida e potente, ma "costosa" perché richiede energia per produrre e conservare gli enzimi in piccoli granuli ed è quindi riservata a minacce concrete già identificate.
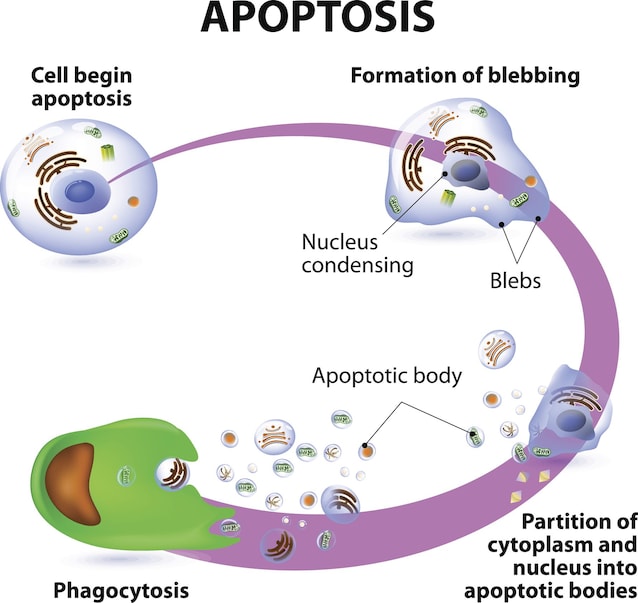
La fase esecutiva: dove tutte le vie convergono
Indipendentemente dal segnale di partenza, tutte e tre le vie convergono sulla stessa fase di esecuzione, dominata dalle cosiddette caspasi effettrici (o esecutrici): caspasi-3, caspasi-6 e caspasi-7, attivate dalle caspasi iniziatrici (caspasi-8 e -9).
Una volta attive, le caspasi effettrici tagliano centinaia di substrati proteici diversi nella cellula, condensano la cromatina (il DNA impacchettato insieme alle proteine istoniche nel nucleo), frammentano il DNA e tutto ciò che c’è nella cellula formando i cosiddetti corpi apoptotici, piccole vescicole delimitate da membrana che vengono rapidamente riconosciute e “mangiate” (fagocitosi) dai macrofagi.
Perché una cellula dovrebbe decidere di autodistruggersi?
Le nostre cellule si dividono continuamente e l’apoptosi è proprio il contrappeso di questa proliferazione cellulare. Se non ci fosse, che ne faremmo di tutte quelle cellule in più? Grazie all’apoptosi è infatti possibile rinnovare ripetutamente i tessuti, il cosiddetto turnover cellulare.
Spesso poi ci dimentichiamo che ogni giorno nelle nostre cellule avvengono centinaia di migliaia di mutazioni: grazie a meccanismi di controllo (come la proteina p53) quando se ne accumulano troppe, la cellula decide di autodistruggersi.
È utile nello sviluppo embrionale, quando i tessuti devono essere ancora “scolpiti”, per esempio per eliminare le membrane tra le dita. Ed è alla base delle mestruazioni, come riporta già un lavoro di Shikone, Yamoto e colleghi pubblicato nel 1996, quando non ricevendo più ormoni, se non c’è stata fecondazione, le cellule dell’endometrio si autodistruggono e provocano la mestruazione. E, ovviamente, permette di identificare e distruggere cellule infette o tumorali.
Come viene riconosciuta la cellula da eliminare?
I recettori di morte sono presenti su tutte le cellule quindi come si fa a capire se deve essere eliminata o meno? Sulla cellula c’è anche una sorta di “etichetta”, in termini tecnici Complesso Maggiore di Istocompatibilità (Major histocompatibility complex, MHC), per gli umani Human leukocyte antigen, HLA. Tutte le cellule hanno un MHC-I sulla propria membrana per dire al sistema immunitario “tranquillo, sono un pezzo dell’organismo” (riconoscimento self).
Quando una cellula è infetta, l’MHC-I si modifica, esponendo, come se fosse una vetrina, pezzi di proteine virali, batteriche o tumorali. I linfociti T citotossici riconoscono che c’è qualcosa di estraneo (non-self), attivano il legame tra ligandi e recettori di morte e di conseguenza la procedura di esecuzione.
Le cellule NK, invece, riconoscono soprattutto le cellule che non hanno o hanno meno MHC-I in superficie ("missing self"), un segnale tipico di infezioni o cellule tumorali, che si credono furbi cancellando le “vetrine” che li potrebbero tradire.
Cosa succede quando c’è uno squilibrio di apoptosi (troppa o troppa poca)
L’importanza di questa strada verso l’abisso delle cellule è palese se pensiamo a cosa succede quando non funziona: una cellula che non si autodistrugge quando è arrivato il momento è un bel problema, come ricorda Favaloro nella sua revisione sul ruolo dell'apoptosi nelle malattie. Per esempio, i tumori sono tristemente conosciuti proprio per la proliferazione cellulare incontrollata accompagnata dalla mancanza di autodistruzione, perché sono in grado di zittire i controllori delle anomalie del DNA (il p53), o di produrre “poliziotti” in eccesso della via intrinseca.
Prasad, in una revisione nel 2003 ricorda l'importanza dell'apoptosi nelle malattie autoimmuni. Se i linfociti T e B autoreattivi, che normalmente vengono eliminati per apoptosi dopo l'attivazione (un processo chiamato "activation-induced cell death"), sopravvivono, possono attaccare i tessuti dell'organismo. Per non parlare di cellule infette con virus e batteri che, se non eliminate, possono continuare a prosperare.
Ma anche un eccesso di apoptosi può creare problemi. Pensiamo alle malattie neurodegenerative come Alzheimer, Parkinson, Huntington, SLA, dove l'accumulo di proteine mal ripiegate contribuisce a innescare la morte apoptotica dei neuroni, cellule che, una volta perse, non vengono sostituite in modo efficace. O anche nell’AIDS, dove a soccombere all’apoptosi incontrollata sono i linfociti T, che dovrebbero difendere l’organismo. Insomma, come sempre, il nostro organismo ha bisogno di equilibrio, anche quando si tratta di vita e di morte.
;Resize,width=578;)
;Resize,width=767;)
;Resize,width=727;)
;Resize,width=727;)
;Resize,width=727;)
;Resize,width=727;)
;Resize,width=727;)

